 Your new post is loading...
Your new post is loading...
Authors: Hikaru Sato and Hisayo Yamane.
Journal of Experimental Botany (2024)
Abstract: "As sessile organisms, plants enter periods of dormancy in response to environmental stresses to ensure continued growth and reproduction in future. During dormancy, plant growth is suppressed, adaptive/survival mechanisms are exerted, and stress tolerance increases over a prolonged period until the plants resume their development or reproduction under favorable conditions. In this review, we focus on seed dormancy and bud dormancy, which are critical for adaptations to fluctuating environmental conditions. We provide an overview of the physiological characteristics of both types of dormancy as well as the importance of the phytohormones abscisic acid and gibberellin for establishing and releasing dormancy, respectively. Additionally, recent epigenetic analyses have revealed that dormancy establishment and release are associated with the removal and deposition of histone modifications at the loci of key regulatory genes influencing phytohormone metabolism and signaling, including DELAY OF GERMINATION 1 and DORMANCY-ASSOCIATED MADS-box genes. We discuss our current understanding of the physiological and molecular mechanisms required to establish and release seed dormancy and bud dormancy, while also describing how environmental conditions control dormancy depth, with a focus on the effects of histone modifications."
Authors: Jihua Ding, Kejing Wang, Shashank Pandey, Mariano Perales, Isabel Allona, Md Rezaul Islam Khan, Victor B. Busov, Rishikesh P. Bhalerao
Journal of Experimental Botany (2024)
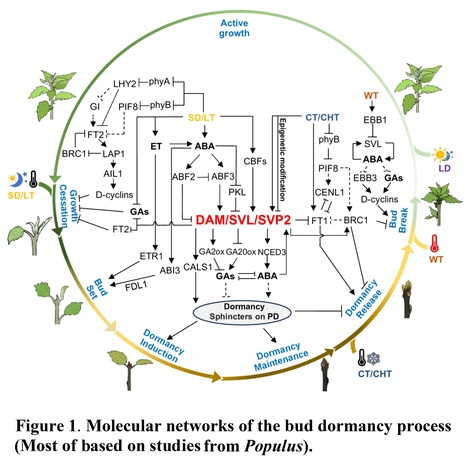
Abstract: "Seasonal bud dormancy in perennial woody plants is a crucial and intricate process that is vital for the survival and development of plants. Over the past few decades, significant advancements have been made in understanding many features of bud dormancy, particularly in model species, where certain molecular mechanisms underlying this process have been elucidated. In this review, we provide an overview of recent molecular progress in understanding bud dormancy in trees, with a specific emphasis on the integration of common signaling and molecular mechanisms identified across different tree species. Additionally, we address some challenges that have emerged in the in-depth understanding of bud dormancy and offer insights for future studies."
Authors: Nora Flynn.
The Plant Cell (2024)
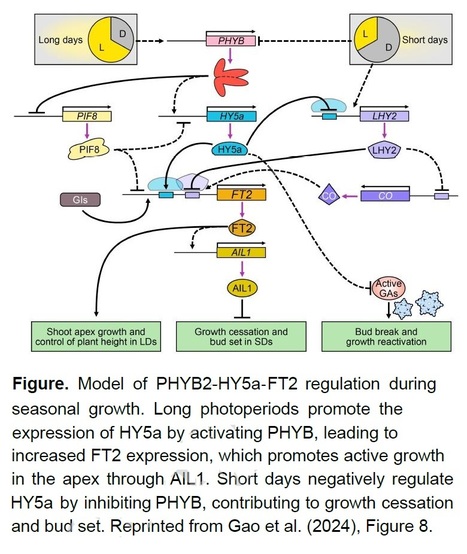
Excerpts: "Yongfeng Gao and colleagues (Gao et al. 2024) expose a new regulatory model for short-day-induced dormancy that involves a poplar (Populus tomentosa) ortholog of the Arabidopsis photomorphogenesis regulatory factor ELONGATED HYPOCOTYL 5 (HY5), HY5a. The model introduces poplar HY5a as a coordinator of growth cessation and bud set that promotes FLOWERING LOCUS T2 (FT2) transcription through both direct and indirect routes (Figure)."
"Finally, the phenotypes of HY5a-OE and HY5a-KO plants indicated that HY5a may also be involved in bud burst. To further investigate this hypothesis, the authors compared the transcript abundances of genes related to the synthesis and breakdown of gibberellins (GA), plant hormones involved in dormancy release (Rinne et al., 2011). The activity of GA synthesis genes increased without HY5a, along with the levels of active GA. Therefore, HY5a is not only a novel regulator of short-day-induced dormancy in poplar, but also a broader controller of seasonal growth that negatively regulates GA in buds. Figure."
Authors: Ya-Lin Zhao, Yong Li, Ke Cao, Jia-Long Yao, Hang-Ling Bie, Irshad Ahmad Khan, Wei-Chao Fang, Chang-Wen Chen, Xin-Wei Wang, Jin-Long Wu, Wen-Wu Guo and Li-Rong Wang.
Plant Physiology (2023)
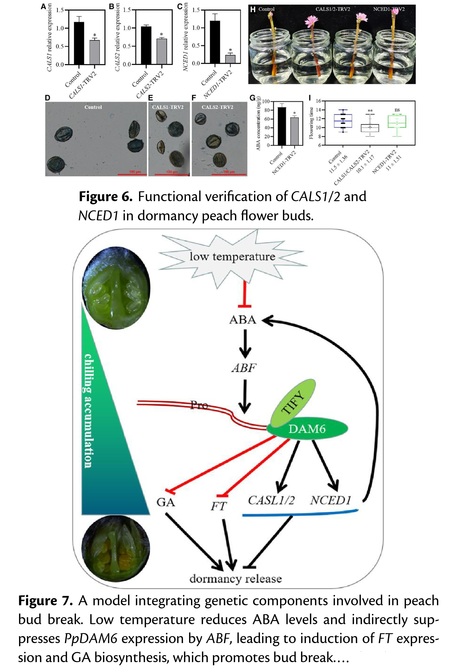
Abstract: "Bud dormancy is crucial for winter survival and is characterized by the inability of the bud meristem to respond to growth-promotive signals before the chilling requirement (CR) is met. However, our understanding of the genetic mechanism regulating CR and bud dormancy remains limited. This study identified PpDAM6 (DORMANCY-ASSOCIATED MADS-box) as a key gene for CR using a genome-wide association study analysis based on structural variations in 345 peach (Prunus persica (L.) Batsch) accessions. The function of PpDAM6 in CR regulation was demonstrated by transiently silencing the gene in peach buds and stably overexpressing the gene in transgenic apple (Malus × domestica) plants. The results showed an evolutionarily conserved function of PpDAM6 in regulating bud dormancy release, followed by vegetative growth and flowering, in peach and apple. The 30-bp deletion in the PpDAM6 promoter was substantially associated with reducing PpDAM6 expression in low-CR accessions. A PCR marker based on the 30-bp indel was developed to distinguish peach plants with non-low and low CR. Modification of the H3K27me3 marker at the PpDAM6 locus showed no apparent change across the dormancy process in low- and non-low- CR cultivars. Additionally, H3K27me3 modification occurred earlier in low-CR cultivars on a genome-wide scale. PpDAM6 could mediate cell–cell communication by inducing the expression of the downstream genes PpNCED1 (9-cis-epoxycarotenoid dioxygenase 1), encoding a key enzyme for ABA biosynthesis, and CALS (CALLOSE SYNTHASE), encoding callose synthase. We shed light on a gene regulatory network formed by PpDAM6-containing complexes that mediate CR underlying dormancy and bud break in peach. A better understanding of the genetic basis for natural variations of CR can help breeders develop cultivars with different CR for growing in different geographical regions."
Authors: HANG Jun-nan, WU Bo-wen, QIU Di-yang, YANG Guo, FANG Zhong-ming and ZHANG Ming-yong.
Journal of Integrative Agriculture (2023)
Abstract: "Low-affinity nitrate transporter gene members have been identified in subfamilies 4–8 of the rice nitrate transporter 1/peptide transporter family (NPF), but the OsNPF3 subfamily responsible for nitrate and phytohormone transport and rice growth and development is unknown. In this study, we described OsNPF3.1 as an essential nitrate and phytohormone transporter gene for rice tillering and nitrogen utilization efficiency (NUtE). OsNPF3.1 possesses four major haplotypes of its promoter sequence in 517 cultivars, and its expression is positively associated with tiller number. Its expression was higher in the basal part, culm, and leaf blade than in other parts, and was strongly induced by nitrate, abscisic acid (ABA) and gibberellin 3 (GA3) in the root and shoot of rice, respectively. Electrophysiological experiment demonstrated that OsNPF3.1 is a pH-dependent low-affinity nitrate transporter, with rice protoplast uptake assays showing it as an ABA and GA3 transporter. OsNPF3.1 over-expression significantly promoted ABA accumulation in the roots and GA accumulation in the basal part to inhibit axillary bud outgrowth and rice tillering, especially at high nitrate concentrations. The NUtE of OsNPF3.1-overexpressing plants was enhanced under low and medium nitrate concentrations, whereas the NUtE of OsNPF3.1 CRISPR plants was increased under high nitrate concentrations. The results indicate that OsNPF3.1 transports nitrate and phytohormones in different rice tissues under different nitrate concentrations. The altered OsNPF3.1 expression improves NUtE through OsNPF3.1-overexpressing and CRISPR lines at low and high nitrate concentrations, respectively."
Author: Sebastian R. Moreno
Plant Physiology (2023)
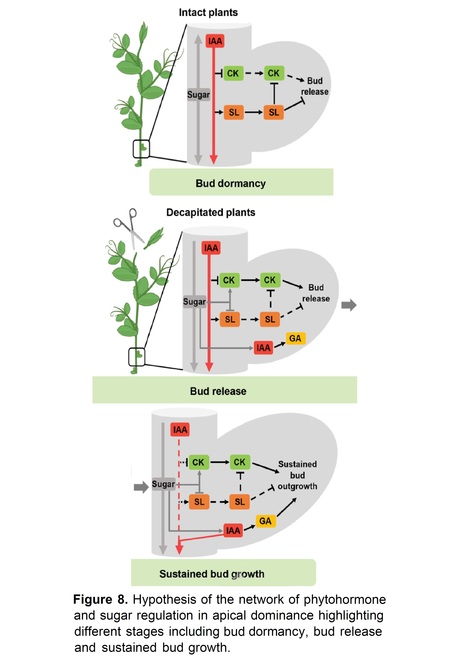
Excerpts: "In this issue of Plant Physiology, Cao et al. (2023) provide evidence supporting the role of sucrose as the signal that triggers changes in CK, auxin, and gibberellin (GA) concentrations to modulate bud outgrowth and to sustain bud growth. The researchers observed that in pea (Pisum sativum) buds outside the auxin-depletion region after decapitation, bud release is triggered by sugar-elicited changes in CK levels with the subsequent downregulation of the inhibitory role of SL (Figure 1).
"Unlike CK, application of SL on axillary buds represses bud outgrowth (Gomez-Roldan et al., 2018). However, the crosstalk between SL and CK behind bud outgrowth is not completely understood. In this paper, through quantifying gene expression of several CK-related genes, the authors nicely demonstrated that SL inhibits CK levels by increasing CK degradation and decreasing CK biosynthesis. In addition, the authors quantified bud growth after treating with synthetic CK BAP and GR24 (synthetic SL) treatments. They found that sucrose and CK treatments could overcome SL-inhibited bud release. Thus, the authors established a very clear connection between SL, CK, and sucrose during early stage of bud growth and provided insights into the mechanism responsible for releasing axillary buds."
Authors: Da Cao, Tinashe Chabikwa, Francois Barbier, Elizabeth A. Dun, Franziska Fichtner, Lili Dong, Stephanie C. Kerr and Christine A. Beveridge.
Plant Physiology (2023)
Short summary: Sugars and cytokinin initiate bud release independently of auxin and suppress inhibition by SL. Afterward, auxin in buds regulates gibberellin to promote sustained bud growth.
Abstract: "The inhibition of shoot branching by the growing shoot tip of plants, termed apical dominance, was originally thought to be mediated by auxin. Recently the importance of the shoot tip sink strength during apical dominance has re-emerged with recent studies highlighting roles for sugars in promoting branching. This raises many unanswered questions on the relative roles of auxin and sugars in apical dominance. Here we show that auxin depletion after decapitation is not always the initial trigger of rapid cytokinin increases in buds that are instead correlated with enhanced sugars. Auxin may also act through strigolactones which have been shown to suppress branching after decapitation, but here we show that strigolactones do not have a significant effect on initial bud outgrowth after decapitation. We report here that when sucrose or cytokinin is abundant, strigolactones are less inhibitory during the bud release stage compared to during later stages and that strigolactone treatment rapidly inhibits cytokinin accumulation in pea (Pisum sativum) axillary buds of intact plants. After initial bud release, we find an important role of gibberellin in promoting sustained bud growth downstream of auxin. We are therefore able to suggest a model of apical dominance that integrates auxin, sucrose, strigolactones, cytokinins and gibberellins and describes differences in signalling across stages of bud release to sustained growth."
Authors: Wenxing Chen, Yosuke Tamada, Hisayo Yamane, Miwako Matsushita, Yutaro Osako, Mei Gao-Takai, Zhengrong Luo and Ryutaro Tao.
The Plant Journal (2022)
Abstract: "Bud dormancy helps woody perennials survive winter and activate robust plant development in the spring. For apple (Malus × domestica), short-term chilling induces bud dormancy in autumn, then prolonged chilling leads to dormancy release and a shift to a quiescent state in winter, with subsequent warm periods promoting bud break in spring. Epigenetic regulation contributes to seasonal responses such as vernalization. However, how histone modifications integrate seasonal cues and internal signals during bud dormancy in woody perennials remains largely unknown. Here, we show that H3K4me3 plays a key role in establishing permissive chromatin states during bud dormancy and bud break in apple. The global changes in gene expression strongly correlated with changes in H3K4me3, but not H3K27me3. High expression of DORMANCY-ASSOCIATED MADS-box (DAM) genes, key regulators of dormancy, in autumn was associated with high H3K4me3 levels. In addition, known DAM/SHORT VEGETATIVE PHASE (SVP) target genes significantly overlapped with H3K4me3-modified genes as bud dormancy progressed. These data suggest that H3K4me3 contributes to the central dormancy circuit, consisting of DAM/SVP and abscisic acid (ABA), in autumn. In winter, the lower expression and H3K4me3 levels at DAMs and gibberellin metabolism genes control chilling-induced release of dormancy. Warming conditions in spring facilitate the expression of genes related to phytohormones, the cell cycle, and cell wall modification by increasing H3K4me3 toward bud break. Our study also revealed that activation of auxin and repression of ABA sensitivity in spring are conditioned at least partly through temperature-mediated epigenetic regulation in winter."
Authors: Feng-Pan Wang, Pan-Pan Zhao, Lei Zhang, Heng Zhai, Muhammad Abid and Yuan-Peng Du.
Frontiers in Plant Science (2022(
Abstract: "Dormancy is a common survival strategy in plants to temporarily suspend visible growth under unsuitable conditions. The elaborate mechanism underlying bud break in perennial woody plants is gradually illustrated. Here, we identified a grape vine WRKY transcription factor, VvWRKY37, which was highly expressed in dormant buds. It was particularly induced by the application of exogenous abscisic acid, and depressed on exposure to gibberellin and low temperature (4°C) stress at the transcript level. The yeast one-hybrid assay confirmed that VvWRKY37 had a transcriptional activity. Ectopic over-expression of VvWRKY37 significantly delayed bud break of transgenic poplar plants. As an ABA-inducible gene, VvWRKY37 also depressed the expression of ABA catabolic gene CYP707As and enhanced the accumulation of endogenous ABA in transgenic poplar plants. The molecular pieces of evidence showed that VvWRKY37 preferentially recognized and bound W-box 5′-G/CATTGACT/C/G-3′ cis-element in vitro. Additionally, VvABI5 and VvABF2 acted as the upstream transcriptional activators of VvWRKY37 via protein-DNA interactions. Taken together, our findings provided valuable insights into a new regulatory mechanism of WRKY TF by which it modulates bud break through ABA-mediated signaling pathways."
Authors: Sen Li, Qingjie Wang, Binbin Wen, Rui Zhang, Xiuli Jing, Wei Xiao, Xiude Chen, Qiuping Tan and Ling Li.
Frontiers in Plant Science (2021)
Abstract: "Gibberellin (GA) plays a key role in the release of bud dormancy and the GA receptor GID1 (GIBBERELLIN INSENSITIVE DWARF1) and DELLA protein are the GA signaling parts, but the molecular mechanism of GA-GID1-DELLA module regulating leaf bud dormancy in peach (Prunus persica) is still not very clear. In this study, we isolated and characterized the GID1 gene PpGID1c from the peach cultivar “Zhong you No.4.” Overexpressing PpGID1c in Arabidopsis promoted seed germination, which indicated that PpGID1c has an important function in dormancy. The expression level of PpGID1c in peach leaf buds during endodormancy release was higher than that during ecodormancy and was positively correlated with GA4 levels. Our study also found that GA4 had the most obvious effect on promoting the bud break, indicating that GA4 may be the key gibberellin to promoting peach leaf bud endodormancy release. Moreover, a quantitative real-time PCR (qRT-PCR) found that GA4 could increase the expression of the gibberellin signaling gene PpDELLA2. A yeast two-hybrid (Y2H) assay suggested that the PpGID1c interaction with the PpDELLA1 protein was not dependent on gibberellin, while the PpGID1c interaction with PpDELLA2 required GA4 or another gibberellin. These findings suggested that the GA4-GID1c-DELLA2 module regulates peach leaf bud endodormancy release, with this finding significantly enhancing our comprehensive understanding of bud endodormancy release and revealing a new mechanism for regulating leaf bud endodormancy release in peach."
Authors: Zhang Yuxi, Yuan Yanchao, Liu Zejun, Zhang Tao, Li Feng, Liu Chunying and Gai Shupeng.
BMC Plant Biology (2021)
Abstract: "Background - Sufficient low temperature accumulation is the key strategy to break bud dormancy and promote subsequent flowering in tree peony anti-season culturing production. Exogenous gibberellins (GAs) could partially replace chilling to accelerate dormancy release, and different kinds of GAs showed inconsistent effects in various plants. To understand the effects of exogenous GA3 and GA4 on dormancy release and subsequent growth, the morphological changes were observed after exogenous GAs applications, the differentially expressed genes (DEGs) were identified, and the contents of endogenous phytohormones, starch and sugar were measured, respectively. Results - Morphological observation and photosynthesis measurements indicated that both GA3 and GA4 applications accelerated bud dormancy release, but GA3 feeding induced faster bud burst, higher shoot and more flowers per plant. Full-length transcriptome of dormant bud was used as the reference genome. Totally 124 110 459, 124 015 148 and 126 239 836 reads by illumina transcriptome sequencing were obtained in mock, GA3 and GA4 groups, respectively. Compared with the mock, there were 879 DEGs and 2 595 DEGs in GA3 and GA4 group, 1 179 DEGs in GA3 vs GA4, and 849 DEGs were common in these comparison groups. The significant enrichment KEGG pathways of 849 DEGs highlighted plant hormone signal transduction, starch and sucrose metabolism, cell cycle, DNA replication, etc. Interestingly, the contents of endogenous GA1, GA3, GA4, GA7 and IAA significantly increased, ABA decreased after GA3 and GA4 treatments by LC–MS/MS. Additionally, the soluble glucose, fructose and trehalose increased after exogenous GAs applications. Compared to GA4 treatment, GA3 induced higher GA1, GA3 and IAA level, more starch degradation to generate more monosaccharide for use, and promoted cell cycle and photosynthesis. Higher expression levels of dormancy-related genes, TFL, FT, EBB1, EBB3 and CYCD, and lower of SVP by GA3 treatment implied more efficiency of GA3. Conclusions - Exogenous GA3 and GA4 significantly accelerated bud dormancy release and subsequent growth by increasing the contents of endogenous bioactive GAs, IAA, and soluble glucose such as fructose and trehalose, and accelerated cell cycle process, accompanied by decreasing ABA contents. GA3 was superior to GA4 in tree peony forcing culture, which might because tree peony was more sensitive to GA3 than GA4, and GA3 had a more effective ability to induce cell division and starch hydrolysis. These results provided the value data for understanding the mechanism of dormancy release in tree peony."
|
Authors: Tao Zhang, Xinyu Wang, Yanchao Yuan, Shoujie Zhu, Chunying Liu, Yuxi Zhang, Shupeng Gai.
Horticulture Research (2024)
Abstract: "Bud endodormancy in perennial plants is a sophisticated system that adapts to seasonal climatic changes. Growth-promoting signals such as low temperature and gibberellins (GAs) are crucial for facilitating budbreak following endodormancy release (EDR). However, the regulatory mechanisms underlying GA-mediated budbreak in tree peony (Paeonia suffruticosa) remain unclear. In tree peony, the expression of PsmiR159b among three differentially expressed miR159 members was inhibited with the prolonged chilling, and overexpression of PsMIR159b delayed budbreak, whereas silencing PsmiR159b promoted budbreak after dormancy. PsMYB65, a downstream transcription factor in the GA pathway, was induced by prolonged chilling and exogenous GA3 treatments. PsMYB65 was identified as a target of PsmiR159b, and promoted budbreak in tree peony. RNA-seq of PsMYB65-silenced buds revealed significant enrichment in the GO terms regulation of ‘cell cycle’ and ‘DNA replication’ among differentially expressed genes. Yeast one-hybrid and electrophoretic mobility shift assays demonstrated that PsMYB65 directly bound to the promoter of the type-D cyclin gene PsCYCD3;1. Dual-luciferase reporter assay indicated that PsMYB65 positively regulate PsCYCD3;1 expression, suggesting that miR159b-PsMYB65 module contributes to budbreak by influencing the cell cycle. Our findings revealed that the PsmiR159b-PsMYB65 module functioned in budbreak after dormancy by regulating cell proliferation, providing valuable insights into the endodormancy release regulation mechanism."
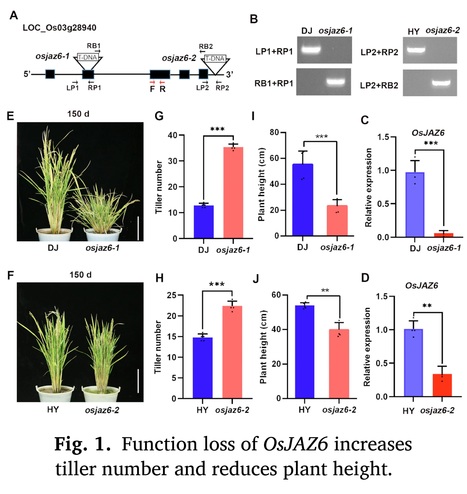
Authors: Wanmin Wang, Zizhao Xie, Yuanyuan Wu, Ying Sun, Chenghang Zhan, Liang Jin and Junli Huang.
Environmental and Experimental Botany (2024)
Highlights: • OsJAZ6 modulates rice tillering and drought response by integrating JA with GA signaling. • OsJAZ6 controls the tiller bud growth but not formation. • OsJAZ6 interacts with SLR1 to promote its degradation, which further destabilizes MOC1. • OsJAZ6 and SLR1 have opposite functions in regulating rice tiller bud growth and drought tolerance.
Abstract: "Jasmonic acid (JA) plays crucial functions during plant growth and stress response, but its roles and regulatory mechanism in plant branching remain largely unknown. Rice basal branching (tillering) is an essential agronomic trait that affects crop production. Here, we report that OsJAZ6, the repressor of JA signaling, negatively modulates rice tillering and drought stress tolerance. Loss-of-function mutants of OsJAZ6 exhibit a significant increase in tiller number, while OsJAZ6ΔJas-overexpression lines produce fewer tillers than wild-type plants. Further investigations show that function loss of OsJAZ6 promotes the tiller bud growth rather than formation. Mechanistic studies show that OsJAZ6 interacts with rice DELLA/SLR1 (SLENDER RICE 1), a transcription repressor of gibberellin (GA) signaling, and the interaction promotes SLR1 degradation, which further facilitates the degradation of rice tillering regulator MOC1 (MONOCULM 1), thereby inhibiting the tiller bud growth. In agreement, the slr1 mutant exhibits fewer tillers than wild type. Consistently, application of JA promotes the growth of tiller bud and thus increases the tiller number, while GA treatment results in opposite result. Meanwhile, osjaz6 mutants display enhanced drought tolerance, coupled with increased JA sensitivity, while the slr1 mutant shows the reverse behavior. Collectively, our data demonstrate that OsJAZ6 negatively modulates rice tillering as well as drought stress tolerance by destabilizing SLR1 protein. Our data shed light on the regulatory mechanism of controlling the tiller development and drought stress response in rice by the JA-OsJAZ6-SLR1 module."
Authors: Yongfeng Gao, Zihao Chen, Qian Feng, Tao Long, Jihua Ding, Peng Shu, Heng Deng, Peizhi Yu, Wenrong Tan, Siqin Liu, Lucas Gutierrez Rodriguez, Lijun Wang, Víctor Resco de Dios and Yinan Yao.
The Plant Cell (2024)
Abstract: "Photoperiod is a crucial environmental cue for phenological responses, including growth cessation and winter dormancy in perennial woody plants. Two regulatory modules within the photoperiod pathway explain bud dormancy induction in poplar (Populus spp.): the circadian oscillator LATE ELONGATED HYPOCOTYL 2 (LHY2) and GIGANTEA-like genes (GIs) both regulate the key target for winter dormancy induction FLOWERING LOCUS T2 (FT2). However, modification of LHY2 and GIs cannot completely prevent growth cessation and bud set under short-day conditions, indicating that additional regulatory modules are likely involved. We identified PtoHY5a, an orthologs of the photomorphogenesis regulatory factor ELONGATED HYPOCOTYL 5 (HY5) in poplar (Populus tomentosa), that directly activates PtoFT2 expression and represses the circadian oscillation of LHY2, indirectly activating PtoFT2 expression. Thus, PtoHY5a suppresses short day-induced growth cessation and bud set. Accordingly, PtoHY5a knockout facilitates dormancy induction. PtoHY5a also inhibits bud-break in poplar by controlling gibberellic acid (GA) levels in apical buds. Additionally, PtoHY5a regulates the photoperiodic control of seasonal growth downstream of phytochrome PHYB2. Thus, PtoHY5a modulates seasonal growth in poplar by regulating the PtoPHYB2–PtoHY5a–PtoFT2 module to determine the onset of winter dormancy, and by fine-tuning GA levels to control bud-break."
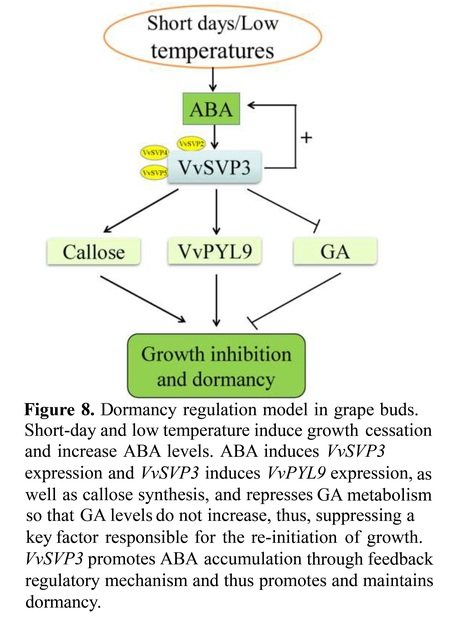
Abstract: Yang Dong, Jing Liu, Liyuan Huang, Lina Yang, Muhammad Khalil-Ur-Rehman, Huan Zheng and Jianmin Tao.
Environmental and Experimental Botany (2023)
Highlights • VvSVP3 induces callose synthesis and VvPYL9 expression, and simultaneously negatively regulates the GA metabolic pathway to maintain dormancy.
Abstract: "Bud dormancy is an essential and complex physiological process in perennial plants. Abscisic acid (ABA) is the major hormone regulating bud dormancy, however, the gene, SHORT VEGETATIVE PHASE (SVP) also plays a pivotal role. The combined regulation of SVP and ABA in the endodormancy network remains poorly characterized. In the present study, we demonstrate the growth inhibitory functions of VvSVP3 by its overexpressing it in tobacco and poplar. VvSVP3-overexpressing poplar plants prematurely stopped growing under short day, exhibiting delayed bud break in early spring. VvSVP3 regulates ABA, the gibberellic acid (GA) pathway, and the callose synthesis to promote dormancy within its network. In addition, exogenous application of ABA positively affects VvSVP3 expression. A transcriptomics and proteomics combined approach identified that VvPYL9 is a target gene of VvSVP3, which directly binds to a CArG motif in the VvPYL9 promoter to activate its expression. In summary, our results revealed the feedback regulation between VvSVP3 gene, ABA metabolism, and the dormancy signaling pathway in grape. It also provides new insights into the functional role of callose and ABA regulation in dormancy."
Authors: Han Dong, Jiachun Wang, Xuewei Song, Chaoyi Hu, Changan Zhu, Ting Sun, Zhiwen Zhou, Zhangjian Hu, Xiaojian Xia , Jie Zhou , Kai Shi, Yanhong Zhou, Christine H. Foyer and Jingquan Yu.
PNAS (2023)
Significance: The orchestration of shoot architecture, which is a major determinant of crop productivity, requires the regulated suppression or activation of -bud outgrowth. We show that light quality regulated tomato bud outgrowth is dependent on the mobile signaling protein HY5 and that HY5 dependent light signaling alone is sufficient to regulate bud outgrowth. HY5 promotes bud growth by direct and brassinosteroid mediated suppression of BRC1. BRC1 blocks the accumulation of cytokinin and gibberellin that regulate bud growth in tomato. In this way, HY5 represses gibberellin mediated stem elongation during photomorphogenesis while activating branching through BRC1 dependent phytohormone regulation. The HY5–BRC1 module thus plays predominant roles in the shoot architecture by orchestrating light quality dependent changes in stem elongation and bud outgrowth.
Abstract: "Light plays an important role in determining plant architecture, which greatly influences crop yield. However, the precise mechanisms by which light signaling regulates bud outgrowth remain to be identified. Here, we show that light regulates bud outgrowth via both HY5 and brassinosteroid (BR)-dependent pathways in tomato. Inactivation of the red-light photoreceptor PHYB, or deficiencies in PHYB or the blue-light photoreceptor CRY1a, inhibits bud outgrowth and leads to decreased accumulation of HY5 protein and increased transcript level of BRANCHED1 (BRC1), a central integrator of branching signals. HY5, functioning as a mobile systemic signal from leaves, promotes bud outgrowth by directly suppressing BRC1 transcript and activating the transcript of BR biosynthesis genes within the lateral buds in tomato. Furthermore, BRC1 prevents the accumulation of cytokinin (CK) and gibberellin (GA) by directly inhibiting the transcript of CK synthesis gene LOG4, while increasing the transcript levels of CK and GA degradation genes (CKX7, GA2ox4, and GA2ox5), leading to an arrest of bud outgrowth. Moreover, bud outgrowth occurs predominantly in the day due to the suppression of BRC1 transcript by HY5. These findings demonstrate that light-inducible HY5 acts as a systemic signaling factor in fine-tuning the bud outgrowth of tomato."
Authors: Manikandan Veerabagu, Christiaan van der Schoot, Veronika Turečková, Danuše Tarkowská, Miroslav Strnad and Päivi L.H. Rinne.
Plant, Cell & Environment (2023)
Abstract: "Perennial para- and endo-dormancy are seasonally separate phenomena. Whereas para-dormancy is the suppression of axillary buds (AXBs) by a growing shoot, endo-dormancy is the short-day elicited arrest of terminal and AXBs. In hybrid aspen (Populus tremula x P. tremuloides) compromising the apex releases para-dormancy, whereas endo-dormancy requires chilling. ABA and GA are implicated in both phenomena. To untangle their roles, we blocked ABA biosynthesis with fluridone (FD), which significantly reduced ABA levels, downregulated GA-deactivation genes, upregulated the major GA3ox-biosynthetic genes, and initiated branching. Comprehensive GA-metabolite analyses suggested that FD treatment shifted GA production to the non-13-hydroxylation pathway, enhancing GA4 function. Applied ABA counteracted FD effects on GA metabolism and downregulated several GA3/4-inducible α- and γ-clade 1,3-β-glucanases that hydrolyse callose at plasmodesmata (PD), thereby enhancing PD-callose accumulation. Remarkably, ABA-deficient plants repressed GA4 biosynthesis and established endo-dormancy like controls but showed increased stress sensitivity. Repression of GA4 biosynthesis involved short-day induced DNA methylation events within the GA3ox2 promoter. In conclusion, the results cast new light on the roles of ABA and GA in dormancy cycling. In para-dormancy, PD-callose turnover is antagonized by ABA, whereas in short-day conditions, lack of GA4 biosynthesis promotes callose deposition that is structurally persistent throughout endo-dormancy."
Authors: Da Cao, François Barbier, Elizabeth A. Dun, Franziska Fichtner, Lili Dong, Stephanie C. Kerr and Christine A Beveridge.
bioRxiv (2022)
Abstract: "The inhibition of shoot branching by the growing shoot tip of plants, termed apical dominance, was originally thought to be mediated by auxin. Recently the importance of the shoot tip sink strength during apical dominance has re-emerged with recent studies highlighting roles for sugars in promoting branching. This raises many unanswered questions on the relative roles of auxin and sugars in apical dominance. Here we show that auxin regulation of cytokinins, which promote branching, is significant only after an initial stage of branching we call bud release. During this early bud release stage, rapid cytokinin increases are associated with enhanced sugars. Auxin may also act through strigolactones which have been shown to suppress branching after decapitation, but here we show that strigolactones do not have a significant effect on initial bud outgrowth after decapitation. We report here that when sucrose or cytokinin is abundant, strigolactones are less inhibitory during the bud release stage compared to later stages and that strigolactone treatment rapidly inhibits cytokinin accumulation in pea axillary buds of intact plants. After initial bud release, we find an important role of gibberellin in promoting sustained bud growth downstream of auxin. We are therefore able to suggest a model of apical dominance that integrates auxin, sucrose, strigolactones, cytokinins and gibberellins and describes differences in signalling across stages of bud release to sustained growth."
Authors: Yuzheng Zhang, Qiuping Tan, Ning Wang, Xiangguang Meng, Huajie He, Binbin Wen, Wei Xiao, Xiude Chen, Dongmei Li, Xiling Fu and Ling Li.
Frontiers in Plant Science (2022)
Abstract: "Bud dormancy, which enables damage from cold temperatures to be avoided during winter and early spring, is an important adaptive mechanism of deciduous fruit trees to cope with seasonal environmental changes and temperate climates. Understanding the regulatory mechanism of bud break in fruit trees is highly important for the artificial control of bud break and the prevention of spring frost damage. However, the molecular mechanism underlying the involvement of MYB TFs during the bud break of peach is still unclear. In this study, we isolated and identified the PpMYB52 (Prupe.5G240000.1) gene from peach; this gene is downregulated in the process of bud break, upregulated in response to ABA and downregulated in response to GA. Overexpression of PpMYB52 suppresses the germination of transgenic tomato seeds. In addition, Y2H, Bimolecular fluorescence complementation (BiFC) assays verified that PpMYB52 interacts with a RING-type E3 ubiquitin ligase, PpMIEL1, which is upregulated during bud break may positively regulate peach bud break by ubiquitination-mediated degradation of PpMYB52. Our findings are the first to characterize the molecular mechanisms underlying the involvement of MYB TFs in peach bud break, increasing awareness of dormancy-related molecules to avoid bud damage in perennial deciduous fruit trees."
Authors: Daniela Gómez-Soto, Isabel Allona and Mariano Perales.
Frontiers in Plant Science (2022)
Abstract: "The adaptation and survival of boreal and temperate perennials relies on the precise demarcation of the growing season. Seasonal growth and development are defined by day length and temperature signals. Under long-day conditions in spring, poplar FLOWERING LOCUS T2 (FT2) systemically induces shoot growth. In contrast, FT2 downregulation induced by autumnal short days triggers growth cessation and bud set. However, the molecular role of FT2 in local and long-range signaling is not entirely understood. In this study, the CRISPR/Cas9 editing tool was used to generate FT2 loss of function lines of hybrid poplar. Results indicate that FT2 is essential to promote shoot apex development and restrict internode elongation under conditions of long days. The application of bioactive gibberellins (GAs) to apical buds in FT2 loss of function lines was able to rescue bud set. Expression analysis of GA sensing and metabolic genes and hormone quantification revealed that FT2 boosts the 13-hydroxylation branch of the GA biosynthesis pathway in the shoot apex. Paclobutrazol treatment of WT leaves led to limited internode growth in the stem elongation zone. In mature leaves, FT2 was found to control the GA 13-hydroxylation pathway by increasing GA2ox1 and reducing GA3ox2 expression, causing reduced GA1 levels. We here show that in poplar, the FT2 signal promotes shoot apex development and restricts internode elongation through the GA 13-hydroxylation pathway."
Authors: Jia Feng, Laichao Cheng, Zhenying Zhu, Feiqi Yu, Cheng Dai, Zhongchi Liu, Wen-Wu Guo, Xiao-Meng Wu and Chunying Kang.
Plant Physiology (2021)
Abstract: "Axillary bud development is a major factor that impacts plant architecture. A runner is an elongated shoot that develops from axillary bud and is frequently used for clonal propagation of strawberry. However, the genetic control underlying runner production is largely unknown. Here, we identified and characterized loss of axillary meristems (lam), an ethyl methanesulfonate-induced mutant of the diploid woodland strawberry (Fragaria vesca) that lacked stamens in flowers and had reduced numbers of branch crowns and runners. The reduced branch crown and runner phenotypes were caused by a failure of axillary meristem initiation. The causative mutation of lam was located in FvH4_3g41310, which encodes a GRAS transcription factor, and was validated by a complementation test. lamCR mutants generated by CRISPR/Cas9 produced flowers without stamens and had fewer runners than the wild-type. LAM was broadly expressed in meristematic tissues. Gibberellic acid (GA) application induced runner outgrowth from the remaining buds in lam, but failed to do so at the empty axils of lam. In contrast, treatment with the GA biosynthesis inhibitor paclobutrazol converted the runners into branch crowns. Moreover, genetic studies indicated that lam is epistatic to suppressor of runnerless (srl), a mutant of FveRGA1 in the GA pathway, during runner formation. Our results demonstrate that LAM is required for stamen and runner formation and acts sequentially with GA from bud initiation to runner outgrowth, providing insights into the molecular regulation of these economically important organs in strawberry."
|




















https://acheterozempic5mg.com/
https://acheterozempic5mg.com/shop/
https://acheterozempic5mg.com/product/acheter-adipex-37-5-mg/
https://acheterozempic5mg.com/product/acheter-mounjaro-en-ligne/
https://acheterozempic5mg.com/product/acheter-ozempic-0-25/
https://acheterozempic5mg.com/product/acheter-ozempic-1mg/
https://acheterozempic5mg.com/product/acheter-ozempic-0-5-mg/
https://acheterozempic5mg.com/product/acheter-ozempic-2mg/
https://acheterozempic5mg.com/product/acheter-saxenda-en-ligne/
https://acheterozempic5mg.com/product/acheter-wegovy-en-ligne/
https://acheterozempic5mg.com/product-category/medicaments-contre-l...
https://acheterozempic5mg.com/product/acheter-de-loxycodone/
https://acheterozempic5mg.com/product/acheter-percocet-en-ligne/
https://acheterozempic5mg.com/product/acheter-morphine-en-ligne/
https://acheterozempic5mg.com/product/acheter-depo-testosterone/
https://acheterozempic5mg.com/product/acheter-vyvanse-en-ligne/
https://acheterozempic5mg.com/product/acheter-zepbound-en-ligne/
https://ozempickopen.com/
https://ozempickopen.com/shop/
https://ozempickopen.com/product-categorie/weight-loss-medications/
https://ozempickopen.com/product/koop-ozempic-0-25mg/
https://ozempickopen.com/product/koop-ozempic-0-5mg/
https://ozempickopen.com/product/koop-ozempic-1mg-online/
https://ozempickopen.com/product/koop-ozempic-2mg-online/
https://ozempickopen.com/product/koop-adipex-online/
https://ozempickopen.com/product/mounjaro-kopen/
https://ozempickopen.com/product/koop-mounjaro-online/
https://ozempickopen.com/product/saxenda-kopen/
https://ozempickopen.com/product/koop-wegovy-online/
https://ozempickopen.com/product/koop-zepbound-online/
https://ozempickopen.com/product/koop-rybelsus-online/
https://ozempickopen.com/product/koop-oxycodon-5mg/
https://ozempickopen.com/product/koop-oxycodon-10-mg/
https://ozempickopen.com/product/oxycodon-kopen-online/
https://ozempickopen.com/product/oxycodon-kopen/
https://ozempickopen.com/product/koop-morfine-online/
https://ozempickopen.com/product/koop-depo-testosterone/
https://ozempickopen.com/product/koop-anadrol-oxymetholone/
https://ozempickopen.com/product/koop-oxymetholone-50-mg/
https://ozempiccomprar1mg.com/
https://ozempiccomprar1mg.com/compra-ahora/
https://ozempiccomprar1mg.com/product/ozempic-comprar-sin-receta/
https://ozempiccomprar1mg.com/product/comprar-ozempic-online/
https://ozempiccomprar1mg.com/product/comprar-ozempic/
https://ozempiccomprar1mg.com/product/comprar-ozempic-2mg/
https://ozempiccomprar1mg.com/product/comprar-mounjaro-sin-receta/
https://ozempiccomprar1mg.com/product/comprar-saxenda-espana/
https://ozempiccomprar1mg.com/product/comprar-wegovy-en-linea/
https://ozempiccomprar1mg.com/product/comprar-actiskenan/
https://ozempiccomprar1mg.com/product/comprar-metadona-en-linea/
https://ozempiccomprar1mg.com/product/comprar-oxicodona-5mg/
https://ozempiccomprar1mg.com/product/comprar-oxycontin-10mg-en-linea/
https://ozempiccomprar1mg.com/product/comprar-depo-testosterone/
https://ozempickopen.com/product/koop-oxycodon-10-mg/
https://ozempickopen.com/product/oxycodon-kopen-online/
https://ozempickopen.com/product/oxycodon-kopen/
https://ozempickopen.com/product/koop-oxycontin-online/
https://ozempickopen.com/product/koop-morfine-online/
https://ozempickopen.com/product/koop-adipex-online/
https://ozempickopen.com/product/koop-depo-testosterone/
https://acquistoozempic.com/
https://acquistoozempic.com/acquista-il-tuo-iniettabile-ozempic/
https://acquistoozempic.com/Prodotto/acquista-ozempic-0-25-mg/
https://acquistoozempic.com/Prodotto/acquisto-ozempic-senza-ricetta/
https://acquistoozempic.com/Prodotto/acquista-ozempic-1mg/
https://acquistoozempic.com/Prodotto/acquisto-ephedrine/
https://acquistoozempic.com/Prodotto/acquista-phentermine-online/
https://acquistoozempic.com/Prodotto/acquista-saxenda-online/
https://acquistoozempic.com/Prodotto/acquisto-wegovy-online/
https://acquistoozempic.com/Prodotto/acquista-mounjaro-5-mg/
https://acquistoozempic.com/Prodotto/acquista-zepbound-2-5-mg/
https://acquistoozempic.com/Prodotto/acquista-zepbound-10-mg/
https://acquistasubutexonline.com/
https://acquistasubutexonline.com/prodotto/
https://acquistasubutexonline.com/prodotto/acquista-ozempic/
https://acquistasubutexonline.com/prodotto/acquista-saxenda-online/
https://acquistasubutexonline.com/prodotto/acquista-wegovy-in-italia/
https://acquistasubutexonline.com/prodotto/acquista-zepbound/
https://acquistasubutexonline.com/prodotto/acquista-subutex-online/
https://acquistasubutexonline.com/prodotto/acquista-methadone-in-it...
https://acquistasubutexonline.com/prodotto/acquista-actiskenan-online/
https://acquistasubutexonline.com/prodotto/acquista-morphine-online/
https://acquistasubutexonline.com/prodotto/acquista-ossicodone-online/
https://acquistasubutexonline.com/prodotto/efedrina-acquisto-online/
https://acquistasubutexonline.com/prodotto/acquista-vyvanse-online/
https://acquistasubutexonline.com/prodotto/acquista-methadone-in-it...
https://acquistasubutexonline.com/prodotto/acquista-adipex-online/
https://acquistasubutexonline.com/prodotto/acquista-victoza-online/
https://acquistasubutexonline.com/prodotto/acquista-qsymia-online/
https://acquistasubutexonline.com/
https://acquistasubutexonline.com/prodotto/
https://acquistasubutexonline.com/prodotto/acquista-saxenda-online/
https://acquistasubutexonline.com/prodotto/acquista-subutex-online/
https://acquistasubutexonline.com/prodotto/acquista-victoza-online/
https://acquistasubutexonline.com/prodotto/acquista-qsymia-online/
https://acquistasubutexonline.com/prodotto/acquista-qsymia-online/
https://acquistasubutexonline.com/prodotto/acquista-duromine-online/
https://acquistasubutexonline.com/prodotto/acquista-methadone-in-it...
https://acquistasubutexonline.com/prodotto/acquista-actiskenan-online/
https://acquistasubutexonline.com/prodotto/acquista-morphine-online/
https://acquistasubutexonline.com/prodotto/acquista-ossicodone-online/
https://acquistasubutexonline.com/prodotto/acquista-dexedrine-online/
https://acquistasubutexonline.com/prodotto/acquista-pillole-analog-...
https://acquistasubutexonline.com/prodotto/acquista-ritalin/
https://acquistasubutexonline.com/prodotto/acquista-vyvanse-online/
https://acquistasubutexonline.com/prodotto/efedrina-acquisto-online/
https://acquistasubutexonline.com/prodotto/acquista-toseina-codeine/
https://acquistasubutexonline.com/prodotto/acquista-adipex-online/
https://acquistasubutexonline.com/prodotto/acquista-belviq-online/
https://acquistasubutexonline.com/prodotto/acquista-duromine-online/
https://acquistasubutexonline.com/prodotto/acquista-ozempic/
https://acquistasubutexonline.com/prodotto/acquista-ossicodone-online/
https://acquistasubutexonline.com/prodotto/acquista-qsymia-online/
https://acquistasubutexonline.com/prodotto/acquista-wegovy-in-italia/
https://acquistasubutexonline.com/prodotto/8311acquista-xenical-onl...
Aeropuerto Adolfo Suárez Madrid-Barajas
@gmail.com
koop ozempic Nederland
koop ozempic voor gewichtsverlies
Koop pillen om af te vallen
koop ozempic 0.25 mg
koop depo testosterone 200mg
koop wegovy
acquista saxenda
acquista subutex online
acquista ozempic
acquista qsymia
koop ozempic Amsterdam
koop ozempic Rotterdam
koop ozempic Eindhoven
mounjaro koop
zepbound koop
rybelsus koop
saxenda koop
wegovy koop
koop oxycodon-10
oxycodon 40mg koop
koop oxycodon
oxycontin koop
morfine koop
adipex koop
koop Depo Testosterone
koop oxymetholone 25 mg
koop oxymetholone 50 mg
acquisto ozempic
ozempic prezzo
acquistare ozempic 0.5mg
acquistare ozempic senza ricetta
acquistare ozempic 1mg
acquistare ephedrine
acquistare phentermine
acquistare saxenda
acquistare wegovy
ozempic comprar 1mg "> acquistare ozempic 1mg
comprar ozempic 1mg
ozempic comprar sin receta
ozempic comprar online
ozempic comprar
comprar ozempic 2mg
comprar mounjaro
comprar wegovy
comprar saxenda
comprar oxycontin-10mg
comprar depo testosterone
acheter ozempic
Acheter Ozempic 0.25 mg
Acheter Ozempic 0.25 mg
Acheter Ozempic 0.5 mg
ozempic perte de poid
ozempic saxenda
ozempic wegovy en ligne
ozempic perte de poid
ozempic oxycodone en ligne
ozempic percocet en ligne
ozempic morphine en ligne
ozempic testosterone en ligne
ozempic vyvanse en ligne
ozempic zepbound en ligne
acquista subutex online
acquista saxenda
acquista subutex
acquista victoza
acquista qsymia
acquista duromine
acquista methadone
acquista actiskenan
acquista morphine
acquista oxicodona
acquista sevredol
acquista dexedrine online
acquista pillole
acquista ritalin
acquista vyvanse
efedrina acquisto
acquista adipex
acquista belviq
acquista duromine
acquista ozempic
acquista qsymia